GREENLAND MAP

PAST PROJECTS
Atlantic Puffin Geolocator and Monitoring Project — 2013 to 2016
Status: Data are being analyzed and results written up.
History, objectives, and previous results of study: The Thule area is the northern extent of the Atlantic Puffin’s breeding range along the west coast of Greenland. In this area a number of colonies are known; however, many are extremely small with only 5–10 individuals, and the largest colonies are on Hakluyt Island, the Witch’s Tit, and in the Carey Islands (up to several hundred birds at the largest colony — Carey Islands). Outside of sporadic colony counts made by visitors to these colonies no information is available, and whether these colonies are increasing, decreasing, or stable in size is unknown. Additionally, the timing of migration and wintering locations of Atlantic Puffins nesting in the Thule area is unknown. The only previous research on Atlantic Puffin migration is from a colony in the United Kingdom, from which adult puffins showed no apparent discernable migratory pattern and individuals wintered around Iceland, southern Greenland, the Mediterranean Sea, and throughout the northeast Atlantic.
In the Thule area from the mid-1990s onward we have collected sporadic information on the Atlantic Puffin colony on the Witch’s Tit. This includes counting the number of adults observed during visits in addition to capturing, sampling, and ringing a limited number of individuals each year. Beginning in 2013 we began a more large scale monitoring project at this colony, with the primary objectives being (1) to annually census the colony and capture, band, and sample as many individuals as possible, and (2) use geolocators to determine where puffins from this colony winter. Results from this study will provide information on areas used by wintering Atlantic Puffins from northwest Greenland and help to determine the current status of Atlantic Puffins in the Thule area. A total of 29 geolocators were deployed on adult Atlantic Puffins on the Witch’s Tit during the 2013 (14) and 2014 (15) field seasons.
In 2014, two geolocators were recovered which provided an entire year of data for one individual and 150 days of data for the second. For the single individual that data were recovered from for the entire migratory period, the puffin traveled up and down the west coast of Greenland during migration but spent the vast majority of the winter in the middle of the North Atlantic, over 1,000 km from the nearest land. This is an area few other seabirds have been shown to utilize, and that previously had not been identified as a winter use area for Atlantic Puffins. In 2015 an additional nine geolocators were recovered, three which were deployed in 2013 and six from 2015. During the 2016 field season while multiple individuals were observed with geolocators, we were unable to recapture any of these individuals.
Objectives for 2020 and beyond: A manuscript was written and submitted in 2019 and is currently being revised. By summer 2020 we should have a new and revised manuscript submitted. No additional geolocators will be deployed and it is highly unlikely any additional attempts will be made to recover any further units.
 |
 |
| Downloading data from a geolocator. | Atlantic Puffin. |
Lapland Longspur Geolocator Study — 2013 to 2015
Status: Data have been analyzed, attempting to publish results from lone individual.
History, objectives, and previous results of study: Although Lapland Longspurs are relatively common and likely increasing in number in northwest Greenland, little to no information is available other than anecdotal observations. At Thule Air Base, starting in 2010, we began a banding program to try and determine annual adult turnover in addition to collecting DNA, stable isotope, and pollutant samples from most individuals captured. To date we have banded over 90 individuals, approximately two-thirds of which have been juveniles. While we have had no band recoveries from juveniles, over 50% of adult males have been re-captured the following year. Where individuals from northwest Greenland winter is currently unknown, although based on ring recoveries from west Greenland it is predicted that the primary wintering area will be in the plains area of southern Canada and the northern USA.
Highlights of Research: During 2013 and 2014 16 adult male longspurs were tagged with geolocators in order to determine where this extreme northern populations winters. In 2014, a single adult male was re-captured from 2013 and the data from the geolocator was successfully downloaded. No additional re-captures of individuals tagged with geolocators occurred.
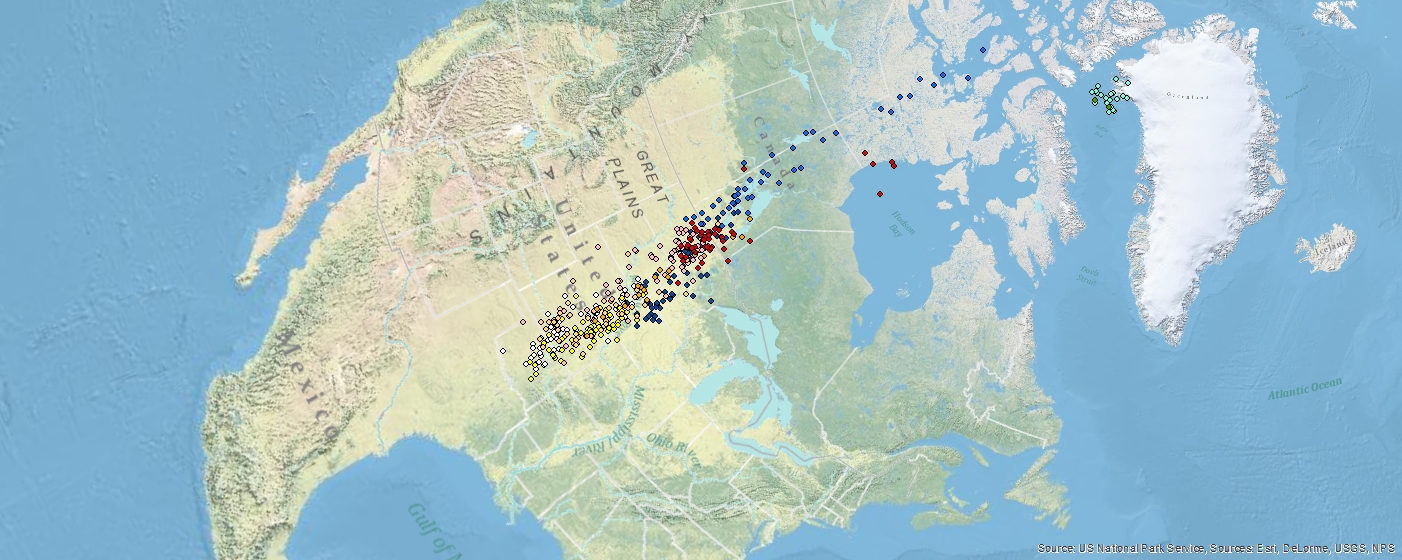 |
|
| August = green, September = light blue, October = blue, November = dark blue, December = white, January = orange, February = light orange, March = dark orange, April = pink, May = red |
Red-necked Phalarope Geolocator Project — 2013 to 2014
Status: No geolocator units recovered, population being monitored.
History, objectives, and previous results of study: Shorebird populations throughout North America appear to have declined over the past several decades. These declines have been documented in both the number of birds observed during migration and on breeding ranges in temperate, sub-Arctic, and Arctic areas. The Red-necked Phalarope breeds in the circumpolar sub-and-low Arctic while migrating to more southern pelagic areas during winter months. Starting in the mid-1980s the number of migrants observed began to severely decline in the North Atlantic. It is not known if this decline is a result of an overall reduction in the total population, a shift in breeding distribution, or a change in migratory route. Within Greenland, Red-necked Phalarope are generally considered common breeders in southern areas and vagrants and very scare breeders in the north. In 2009, while conducting surveys for nesting waterfowl in northwest Greenland, K. Burnham discovered a previously unknown population of nesting Red-necked Phalarope in the Booth Sound area (76.91°N, 70.73°W). Based on historic information from Finn Salomonsen for this area, it is likely that this is a relatively new and likely increasing population. It is unknown where this Greenland breeding population winters. Current known wintering areas for other Red-necked Phalarope populations include the Arabian Sea, East Indies, southern Philippines, northern New Guinea, and the Humboldt Current off of Peru. While on winter grounds they spend their entire time at sea, frequently congregating at areas where plankton are concentrated and lifted to the surface (e.g., upwellings, thermal gradients, etc.), with approximately 2 million individuals observed during migration at an upwelling near Mount Desert Rock off of Maine (USA). In the Atlantic, large wintering areas have yet to be discovered, and birds nesting in the North Atlantic (e.g., Labrador, Greenland) may well winter at a yet to be discovered location in the south Atlantic, cross North America to the Pacific, or travel to southern Europe or Africa.
To determine the wintering location of Red-necked Phalarope that breed in the Thule area, during the 2013 and 2014 field seasons we tagged a total of 14 breeding adults in the Booth Sound area with geolocators. Based on broader surveys of the Booth Sound area from 2010-12, our tagging focused along an approximately 1.5 km long by 0.5 km wide stream/wetland area where over 95% of our total observations occurred. Aside from those individuals tagged with geolocators, an additional 10 and 15 additional adults were observed in 2013 and in 2014, a further six adults were tagged with solely color bands. It was our intention that these additional six would serve as control individuals, to better help us determine what affect, if any, the geolocators were having on survival. During the 2016 field season the wetland area was re-surveyed and no tagged individuals were observed.
 |
|
| Red-necked Phalarope. | Red-necked Phalarope about to be released. |

Black-legged Kittiwake Geolocator Project — 2011 to 2014
Status: Data are being analyzed and written up.
History, objectives, and previous results of study: In 2011, 35 Black-legged Kittiwakes were tagged in the Thule area with geolocators (see above section for what geolocators are) to determine the timing and route of migration along with wintering areas. Previous research (Frederiksen et al. 2011) from northwest Atlantic kittiwake colonies to the south of Thule, and from European colonies, has shown the region in the West Atlantic betweenNewfoundland and the Mid-Atlantic Ridge to have the largest concentration of wintering Black-legged Kittiwakes. While kittiwakes from Thule may very well winter in this same general region, the Thule population is at the extreme northern range for this species, with data on timing of migration and wintering areas for this large population unknown.
Highlights of Research: Placed 35 geolocators on adult Black-legged Kittiwakes on Saunders Island in the Thule area in 2011. In 2012, 17 individuals were re-captured with additional individuals re-captured in 2013 and 2014. Data are currently being analyzed. See the HAI Facebook page for a map showing preliminary results.
 |
 |
| Sampling and placing a geolocator on an adult Black-legged Kittiwake. Hard hats are necessary as many rocks are kicked off the cliff above by the many tens-of-thousands of nesting sea birds. | A pair of Black-legged Kittiwakes on their nest. |
Arctic Tern Geolocator Project — 2010 to 2013
Status: Data have been analyzed, attempting to publish results from single individual.
History, objectives, and previous results of study: Research by Egevang et al. (2010) documented Arctic Terns nesting on Sand Island (74.75° N), east Greenland, to have the longest migration of any species on Earth. Some adults from east Greenland traveled over 80,000 kilometers each year, over wintering in Antarctica. In 2010, High Arctic Institute scientists initiated a similar study in the Thule area, tagging 23 adult Arctic Terns with geolocators to document their year-round migration. Geolocators are small 1–2 gram units that have an internal clock and light sensor which measure when the sun sets and rises. Using this information, two locations can be calculated for each individual day; however, locations can not be calculated when tagged birds are in areas with continuous 24hr sunlight or darkness. Terns in Thule were tagged at a colony at approximately 76.9° N. In 2011, 2012, and 2013 researchers returned to the colony to attempt to re-capture tagged individuals and remove the geolocators. However, in 2011 due to the late departure of sea ice, an ice-bridge remained between the island and mainland for a period of time, allowing Arctic Foxes access to the island. Almost all nests on the island were destroyed or predated, with only three tern nests (that nested very late in the year, after the ice-bridge and fox were gone) found in 2011. Of the adults at the three nests in 2011, one was tagged with a geolocator. This individual was captured and the geolocator removed. No reproduction occurred in 2012 and 2013 and no individuals were observed with geolocators.
Highlights of Research: Placed 23 geolocators on adult Arctic Terns in 2010. Data from a single adult re-captured in 2011 showed it followed air currents both during outward and return migration, similar to results obtained by Egevang et al. (2010) for east Greenland.
 |
 |
| A geolocator is first attached to a band with a wire tie and then the band is put on the Arctic Tern. | Adult Arctic Tern with geolocator. |
Thule Area Nesting Common Eider Survey — 2009
Status: Results Published
Abstract from Burnham et al. 2012
ABSTRACT: Common eider (Somateria mollissima) populations in Greenland severely declined throughout the 20th century. As a result, in 2001, harvest regulations were changed and the length of the hunting season was reduced. Recent data suggest that these changes have been successful, and population regrowth is occurring. In the Avanersuaq District, northwest Greenland, only one systematic survey quantifying the number of nesting eiders had previously been conducted, in 1997 and 1998. Although this district had historically been identified as having the largest number of breeding eiders in Greenland, the 1997 – 98 survey results showed a relatively small estimated population of 5000 pairs. However, it is not known to what extent changes in hunting regulations have affected nesting abundance in this area. Therefore, the Avanersuaq District was system– atically resurveyed during the 2009 breeding season, approximately 11 years after the previous survey. These results showed that the population had increased to 5.4 times its 1997 – 98 size, with an annual compounded growth rate of 15.3%. On a single island, nearly 4500 active nests were observed. Five islands had more than 2600 nests each and comprised 75% of the total nests counted. Along with historical information to account for additional nesting habitat not surveyed, the observed population growth rate from this study suggests that the overall Avanersuaq common eider breeding population size ranges from 25 000 to 30 000 pairs, or roughly half of the total estimated West Greenland breeding population. Despite the significance of the Avanersuaq District as a breeding area for common eiders, we have only limited information about this population. The effects of recent extensions of the hunting season on this
population are also unknown, and the only wintering location information is based on a few individuals banded in the 1920s and 1940s. Additional research on migratory movements is suggested before any further changes are made to hunting regulations.
Thule Area Waterfowl Surveys — 2008 to 2009
Status: Results Published
Abstract from Burnham et al. 2014
ABSTRACT: Breeding populations of Nearctic and Palearctic waterfowl have undergone significant changes in abundance and distribution over the past 50 years. The Avanersuaq District in northwest Greenland is home to an assemblage of waterfowl from both geographic areas; however, minimal historic or current information is avail– able on species abundance. In 2008 and 2009, we conducted field surveys in Greenland from 76.00° to 77.35°N for breeding and non-breeding waterfowl and have collected anecdotal field notes of avian observations over a 20-year period (1993–2012). During these periods, we documented the first observation of a Ross’s goose (Chen rossii) and the first confirmed breeding by lesser snow geese (Chen caerulescens caerulescens) in Greenland.
Northern pintails (Anas acuta) were observed for the first time in northwest Greenland, and a previously unknown breeding location for brent geese (Branta bernicla hrota) was also identified. Local populations of greater snow (C. c.) and Canada geese (B. canadensis) have increased in size. The Booth Sound and Drown Bay wetland areas and many islands throughout the Avanersuaq District were identified as critical habitat for both breeding and non-breeding waterfowl. Further increases in waterfowl abundance, including more frequent rare and new visitors, are likely in the study area as breeding populations further south continue to increase and an ameliorating climate allows for a longer breeding season. These results will prove useful as a baseline for comparisons with future surveys.
Carey Island Bird Survey — 2008
Status: Results Published
Abstract from Burnham & Burnham 2010
ABSTRACT: The Carey Islands of northwest Greenland are infrequently visited by ornithological researchers, and baseline population data for many avian species is limited or absent. This paper highlights the results of a three-day expedition to circumnavigate the largest islands and islets of the Carey Island group to document the population and breeding status of all avian species. Sixteen species were observed, with ten documented breeding. Of particular interest to the avian record of the region is the first documented observation of White-fronted Geese on the Carey Islands and the large number of breeding of Brant Geese.
Gyrfalcon Plumage Color Study — 1998 to 2006
Status: Results Published
Abstract from Johnson & Burnham 2013
ABSTRACT: Plumage colour variation exists among Gyrfalcons throughout their Arctic and sub-Arctic circumpolar distribution, ranging from white through silver and grey to almost black. Although different colour variants coexist within many populations, a few geographical regions, such as northern Greenland, possess a single variant, suggesting that local environments may influence plumage colour variation. In central-west Greenland (66.5–67.5°N), where multiple colour variants exist, white male Gyrfalcons fathered significantly earlier clutches than grey males. No significant association was observed between female colour and lay date. However, significantly more offspring were produced by both male and female white Gyrfalcons than by grey variants when controlling for lay date, and silver Gyrfalcons produced an intermediate number of offspring for both sexes. This pattern was
further supported by breeding plumage colour pairings. Grey females paired with grey males nested significantly later in the season and produced fewer offspring than those paired with white males, whereas no difference in lay date or offspring number was found between white males paired with white or with grey females. The difference in the number of offspring produced at each nest-site was also inversely correlated with the distance to the nearest neighbouring nest, and grey males nested in closer proximity to other nests compared with white and silver colour variants. These results suggest that factors associated with territory occupancy and timing of breeding may regulate reproductive success differently between colour variants, with directional selection favouring
light-coloured Gyrfalcons and resulting in earlier lay date and a high frequency of white plumage colour variants in this population. Although gene flow exists between our study population and those further north (>75°N), white Gyrfalcons prevail where the breeding season duration is even shorter, suggesting that nesting chronology in combination with genetic drift may play an important role in influencing plumage colour polymorphism among Gyrfalcon populations.
Thule Area Peregrine Falcon Study — from 1993 to 2005
Status: Results Published
Abstract from Burnham et al. 2012
ABSTRACT: Peregrine Falcons, Falco peregrinus tundrius, were historically unknown to Inuit and early explorers in the Pituffik (Thule) area, northwest Greenland (75.90–77.60° N). Here we provide information collected from 1993–2005 on what we believe is a recently established and expanding population of High Arctic nesting Peregrines in the area associated with climate change. From 1979 to 2005, the average of the mean monthly temperature, minimum monthly temperature, and maximum monthly temperature for the five-month period, May through September, increased 1.1, 0.5, and 1.6 °c, respectively. Forty-one breeding attempts were recorded at six sites from 1993 to 2005 for this new population. satellite transmitters were used to determine the home ranges and seasonal movements of female Peregrines, with adults traveling an average of 10,794 km at a rate of 205 km/ day on outward migration. During outward migration, the maximum distance traveled by any female on one day was 1,349 km with the maximum total outward and return migrations for single individuals 12,438 and 11,071 km, respectively, to and from south America.
Comparisons with Peregrine populations in Greenland at 67° N and 60.5° N, approximately 1,100 (Kangerlussuaq) and 1,700 (south Greenland) km south of the Pituffik area, respectively, show differences in various aspects of ecology. Based on a lack of both morphological and genetic differences it appears the Pituffik area population is likely a result of the extension of more southern breeding Peregrines moving north and taking advantage of an ameliorating climate and lengthened breeding window. Should climatic amelioration continue, the species may eventually expand its range into the very northernmost land area, Peary Land.
Carbon Dating of Gyrfalcon Nests in Greenland — 2002 to 2004
Status: Results Published
Abstract from Burnham et al. 2009
ABSTRACT: Gyrfalcons Falco rusticolus use the same nest-sites over long periods of time, and in the cold dry climate of Greenland, guano and other nest debris decay slowly. Nineteen guano samples and three feathers were collected from 13 Gyrfalcon nests with stratified faecal accumulation in central-west and northwest Greenland. Samples were 14C dated, with the oldest guano sample
dating to c. 2740–2360 calendar years (cal yr) before present (BP) and three others were probably > 1000 cal yr BP. Feather samples ranged from 670 to 60 cal yr BP. Although the estimated age of material was correlated with sample depth, both sample depth and guano thickness gave a much less reliable prediction of sample age than use of radiocarbon dating on which the margin of error was less. Older samples were obtained from sites farther from the current Greenland Ice Sheet and at higher elevations, while younger samples were closer to the current ice sheet and at lower elevations. Values for d13C showed that Gyrfalcons nesting farther from the Greenland Ice Sheet had a more marine diet, whereas those nesting closer to the ice sheet (= further inland) fed on a more terrestrial diet. The duration of nest-site use by Gyrfalcons is a probable indicator of both the time at which colonization occurred and the palaeoenvironmental conditions and patterns of glacial retreat. Nowhere before has such extreme long-term to present use of raptor nest-sites been documented.
Genetic Structure Among Circumpolar Gyrfalcon Populations — 2001 to 2004
Status: Results Published
Abstract from Johnson et al. 2007
ABSTRACT: Little is known about the possible influence that past glacial events have had on the phylogeography and population structure of avian predators in the Arctic and sub-Arctic. In this study, we use microsatellite and mitochondrial control region DNA variation to investigate the population genetic structure of gyrfalcons (Falco rusticolus) throughout a large portion of their circumpolar distribution. In most locations sampled, the mtDNA data revealed little geographic structure; however, five out of eight mtDNA haplotypes were unique to a particular geographic area (Greenland, Iceland, or Alaska) and the Iceland population differed from others based on haplotype frequency differences (FST). With the microsatellite results, significant population structure (FST, principal components analysis, and cluster analysis) was observed identifying Greenland and Iceland as separate populations, while Norway, Alaska and Canada were identified as a single population consistent with contemporary gene flow across Russia. Within Greenland, differing levels of gene flow between western and eastern sampling locations was indicated with apparent asymmetric dispersal in western Greenland from north to south. This dispersal bias is in agreement with the distribution of plumage colour variants with white gyrfalcons in much higher proportion in northern Greenland. Lastly, because the mtDNA control region sequence differed by only one to four nucleotides from a common haplotype among all gyrfalcons, we infer that the observed microsatellite population genetic structure has developed since the last glacial maximum. This conclusion is further supported by our finding that a closely related species, the saker falcon (Falco cherrug), has greater genetic heterogeneity, including mtDNA haplotypes differing by 1–16 nucleotide substitutions from a common gyrfalcon haplotype. This is consistent with gyrfalcons having expanded rapidly from a single glacial-age refu– gium to their current circumpolar distribution. Additional sampling of gyrfalcons from Fennoscandia and Russia throughout Siberia is necessary to test putative gene flow between Norway and Alaska and Canada as suggested by this study.
The Genetics of Plumage Color in Gyrfalcons — 2001 to 2004
Status: Results Published
Abstract from Johnson et al. 2012
ABSTRACT: Genetic variation at the melanocortin-1 receptor (MC1R) gene is correlated with melanin color variation in a few reported vertebrates. In Gyrfalcon (Falco rusticolus), plumage color variation exists throughout their arctic and subarctic circumpolar distribution, from white to gray and almost black. Multiple color variants do exist within the majority of populations; however, a few areas (e.g., northern Greenland and Iceland) possess a single color variant. Here, we show that the white/ melanic color
pattern observed in Gyrfalcons is explained by allelic variation at MC1R. Six nucleotide substitutions in MC1R resulted in 9 alleles that differed in geographic frequency with at least 2 MC1R alleles observed in almost all sampled populations in Greenland, Iceland, Canada, and Alaska. In north Greenland, where white Gyrfalcons predominate, a single MC1R allele was observed at high
frequency (.98%), whereas in Iceland, where only gray Gyrfalcons are known to breed, 7 alleles were observed. Of the 6 nucleotide substitutions, 3 resulted in amino acid substitutions, one of which (Val128Ile) was perfectly associated with the white/melanic polymorphism. Furthermore, the degree of melanism was correlated with number of MC1R variant alleles, with silver Gyrfalcons
all heterozygous and the majority of dark gray individuals homozygous (Ile128 ). These results provide strong support that MC1R is associated with plumage color in this species.
Genetic Comparison of North American Arctic Peregrine Populations — 2001 to 2004
Status: Results Published
Abstract from Johnson et al. 2010
ABSTRACT:
Background: Our ability to monitor populations or species that were once threatened or endangered and in the process of recovery is enhanced by using genetic methods to assess overall population stability and size over time. This can be
accomplished most directly by obtaining genetic measures from temporally-spaced samples that reflect the overall stability of the population as given by changes in genetic diversity levels (allelic richness and heterozygosity), degree of population differentiation (FST and DEST), and effective population size (Ne). The primary goal of any recovery effort is to produce a longterm self-sustaining population, and these genetic measures provide a metric by which we can gauge our progress and help make important management decisions.
Methodology/Principal Findings: The peregrine falcon in North America (Falco peregrinus tundrius and anatum) was delisted in 1994 and 1999, respectively, and its abundance will be monitored by the species Recovery Team every three years until 2015. Although the United States Fish and Wildlife Service makes a distinction between tundrius and anatum subspecies, our genetic results based on eleven microsatellite loci suggest limited differentiation that can be attributed to an isolation by distance relationship and warrant no delineation of these two subspecies in its northern latitudinal distribution from Alaska through Canada into Greenland. Using temporal samples collected at Padre Island, Texas during migration (seven temporal time periods between 1985–2007), no significant differences in genetic diversity or significant population differentiation in allele frequencies between time periods were observed and were indistinguishable from those obtained from tundrius/anatum breeding locations throughout their northern distribution. Estimates of harmonic mean Ne were variable and imprecise, but always greater than 500 when employing multiple temporal genetic methods.
Conclusions/Significance: These results, including those from simulations to assess the power of each method to estimate
Ne, suggest a stable or growing population, which is consistent with ongoing field-based monitoring surveys. Therefore,
historic and continuing efforts to prevent the extinction of the peregrine falcon in North America appear successful with no
indication of recent decline, at least from the northern latitude range-wide perspective. The results also further highlight the
importance of archiving samples and their use for continual assessment of population recovery and long-term viability.
Migratory Movements of Gyrfalcons in Greenland — 2000 to 2004
Status: Results Published
Abstract from Burnham & Newton 2011
ABSTRACT: Little information exists on the movements of Gyrfalcons Falco rusticolus outside the breeding season, particularly amongst High Arctic populations, with almost all current knowledge based on Low Arctic populations. This study is the first to provide data on summer and winter ranges and migration distances. We highlight a behaviour previously unknown in Gyrfalcons, in which birds winter on sea ice far from land. During 2000–2004, data were collected from 48 Gyrfalcons tagged with satellite transmitters in three parts of Greenland: Thule (northwest), Kangerlussuaq (central-west) and Scoresbysund (central-east). Breeding home-range size for seven adult females varied from 140 to 1197 km2 and was 489 and 503 km2 for two adult males. Complete outward migrations from breeding to wintering areas were recorded for three individuals: an adult male which travelled 3137 km over a 38-day period (83 km ⁄ day) from northern Ellesmere Island to southern Greenland, an adult female which travelled 4234 km from Thule to southern Greenland (via eastern Canada) over an 83-day period (51 km ⁄ day), and an adult female which travelled 391 km from Kangerlussuaq to southern Greenland over a 13-day period (30 km ⁄ day). Significant differences were found in winter home-range size between Falcons tagged on the west coast (383‑6657 km2) and east coast (26 810–63 647 km2). Several Falcons had no obvious winter home-ranges and travelled continually during the non-breeding period, at times spending up to 40 consecutive days at sea, presumably resting on icebergs and feeding on seabirds. During the winter, one juvenile female travelled over 4548 km over an approximately 200-day period, spending over half that time over the ocean between Greenland and Iceland. These are some of the largest winter home-ranges ever documented in raptors and provide the first documentation of the long-term use of pelagic habitats by any falcon. In general, return migrations were faster than outward ones. This study highlights the importance of sea ice and fjord regions in southwest Greenland as winter habitat for Gyrfalcons, and provides the first detailed insights into the complex and highly variable movement patterns of the species.
Bird Survey of the Uummannaq District, west Greenland — 2000
Status: Results Publsihed
Abstract from Burnham et a. 2005
ABSTRACT: From 1905 to 1920 Alfred Bertelsen documented 210 avian breeding sites for 32 species in the Uummannaq District, Greenland, between 70º03’ and 72º03’ N and 50º20’ and 55º40’ W, a land area of about 12 000 km2. In 2000 we re-surveyed 207 of those sites and the remainder of the coastal area, providing results for 25 species, our aim being to determine presence/absence and density of coastal nesting species. For 10 species reported by Bertelsen as common and with quantified numbers (breeding sites and population) at his locations, we found the number of occupied sites was the same for one species (Northern Fulmar Fulmarus glacialis) while nine others had declined. Assuming some species may have relocated breeding sites over time and including all locations where we found those species, when compared to Bertelsen’s results, the number of sites remained the same for two species and declined for eight. Comparing the total observed population numbers (birds present) between Bertelsen’s and our 2000 survey, one species remained about the same or slightly increased (Northern Fulmar) and nine decreased. The species with the most dramatic declines were the Thick-billed Murre Uria lomvia (from 8 sites and over 500 000 pairs to zero), Black-legged Kittiwake Rissa tridactyla (27 sites and 268 000 birds to 7 sites and c. 1100 birds), Razorbill Alca torda (17 sites to 3), Common Eider Somateria mollissima (26 sites to 16), and Gyrfalcon Falco rusticolus (28 sites to 7). The Great Cormorant Phalacrocorax carbo and Great Black-backed Gull Larus marinus are the only species we could determine with certainty that had increased in number of breeding sites (1 to 12 and 0 to 4 sites, respectively). Population declines appear to be a combination of human persecution and human-caused reduction in prey and habitat quality. Unless further conservation measures are taken, continued avian declines are probable.